
Last decades, H. circularisquama has been formed
"red tide" in coastal area of Japan when the water temperature and salinity
became optimal for their growth. Since cell density of H. circularisquama
reached >106 cells/L, the seawater was discolored to dark brown
(coloration similar to "stained iron" or "cocoa"). Left figure shows the
photograph of H. circularisquama red tide occurred in Ago Bay, Mie
Prefecture, in 1992. All discolored water is due to intensive assemblage
of H. circularisquama cells (more than 20 million cells/L). Duration
of H. circularisquama red tide period ranges from 1 week to a few
months. In a case of 1992 red tide occurred in Ago Bay, H. circularisquama
bloom persisted about three month (Matsuyama et al. 1995).
2)Red tide occurrence in western Japan

In Japanese coast, H. circularisquama has appeared
in 1988 in Uranouchi Bay, Kouchi Prefecture at first time and then rapidly
expanded over the western part of Japan. The red tides due to H. circularisquama
has been associated with catastrophic damages on shellfishes aquaculture
in almost regions (Yamamoto and Tanaka 1989, Matsuyama et al. 1996, Yoshida
and Miyamoto 1995). Incidences of this red tide have been increasing recently,
and the economic loss to aquaculture have caused industrial and social
concern (Matsuyama et al. 1996).
Right figure shows the distribution of H. circularisquama
red tide in Japanese coast. H. circularisquama red tide intensively
distributes western Japan, especially, South coast of Mie, from western
Seto Inland Sea to western coast of Kyushu island. The most frequent area
is Ago Bay, where is occurs H. circularisquama red tide from 1992
to 1997. Almost locations where H. circularisquama had bloomed previously,
are along with the major warm currents, so called Kuroshio and Tsushima
Current.
In addition, H. circularisquama red tide areas
are likely overlapped to extensive mariculture site of shellfish, i. e.
Ago Bay (pearl oyser), Hiroshima Bay (oyster), Kumamoto and Suo-nada (short-necked
clam). In recent, simulated experiments suggested that H. circularisquama
can easily immigrate to geologically distance area in association with
pearl oyster transportation (Honjo et al. 1998). Further work is necessary
to larify possibility of artificial dispersal of H. circularisquama
cells by means of human activities.
3)Annual pattern of occurrences and growth profile
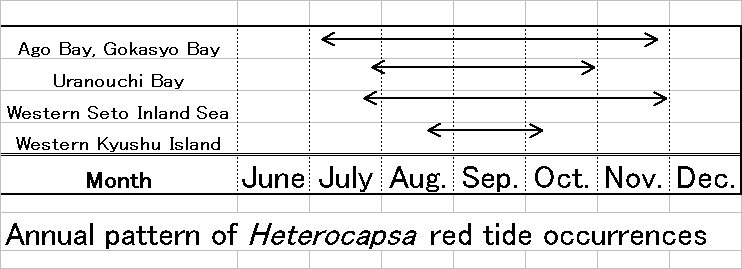
The red tide of H. circularisquama occurs from July to November in embayment of western Japan. Generally, intensive H. circularisquama red tide frequently appears during high water temperature (>23℃) and salinity (>30psu) condition. These field scientific efforts suggested that H. circularisquama likely prefers the comparatively high water and high salinity conditions. According to the laboratory culture experiments, optimal growth of H. circularisquama was demonstrated in combinations with water temperature at >30℃ and salinity at 30 psu, respectively (Yamaguchi et al. 1997). These field growth profile is consistent with those obtained in laboratory culture experiment (Yamaguchi et al. 1997). The optimal growth of representative harmful algae (Alexandrium, Chattonella, Gymnodinium, Heterosigma) which had been bloomed in Japanese coast has demonstrated in combinations with water temperature from 20 to 25゚C, and salinity 20-25 psu, respectively. In western Japan, water temperature seldom exceeds 30 ℃. Thus, growth profile of H. circularisquama, especially optimal growth at 30℃, is greatly different from those of previous harmful algae appears in Japanese coast. Considering these physiological characters, H. circularisquama is more suitable for tropical and sub-tropical zone rather than temperate zone. There have been no records of H. circularisquama red tide and related shellfish mortality in Japan before 1988. It has been argued that whether H. circularisquama is a native species in Japan or not, because no report of H. circularisquama distribution are exsisted in elsewhere except for Japanese Coast. However, though growth response for water temperature is the most genetically stable character, there is possibility that H. circularisquama probably originated from tropical and/or subtropical region and recently invaded to Japanese coast.
4)Water quality changing
To our knowledge, few data available for the relationships
between water quality and recent H. circularisquama proliferation
in Japanese coast. Since horizontal distribution pattern of H. circularisquama
is closely related to those of representative harmful algae Chattonella
and Gymnodinium, eutrofication of coastal area due to effluent loading
and aquaculture development probably underlying in this trend. Meanwhile,
though H. circularisquama sometime blooms in non-eutrophicated area,
this dinoflagellate having highly abilities to oligotrophic condition.
In last decade, selective regulation of phosphorus on
land effluent due to administrative efforts brought about change of nutrient
loading balance such as N:P ratio. These water improvements also highly
likely alter the N:P ratio in Japanese coastal water that maybe stimulate
the novel harmful algal blooms. However, further scientific effort is necessary
to elucidate the relationships between the proliferation due to novel algal
bloom and water qualities.
5)Long-term pattern of red tide occurrences

Historical change of H. circularisquama red tide are shown in above figure. The red tide incidence due to H. circularisquama has been rapidly increasing since early 1990. Why can H. circularisquama inhabit in Japanese coast? Laboratory experiment reveal that H. circularisquama could not grown at 10℃ (Yamaguchi et al. 1997), and does not form typical resting cyst that can overwinter in bottom sediment. The overwintering motile cells, thus, is likely only seed population for the bloom initiations. This eco-physiolocical strategy is greatly disadvantage for the habitation around the temperate and frigid zone. However, the winter water temperature in Japanese coast, has been significantly increasing since late 1980 (e.g. Kodama et al. 1995), its probably due to global climate change (green house effects and El nino). In Seto Inland Sea, for instance, minimum water temperature increased at least 1℃ in last decade, except for 1995-1996 winter. Accordingly, tropical marine animals i. e. green shell mussel Perna viridis, frequently appeared in Seto Inland Sea since early 1990. This hydrographic change is highly likely enhance the overwintering cells of H. circularisquama, that allowed to inhabit in Japanese coast. The relationships between winter water temperature and initial occurrences of nuisance red tide dinoflagellate Gymnodinium mikimotoi is also denoted in previous study (Honjo et al. 1991). Hydrographic change of water temperature brought about unusual sequence of phytoplankton communities and marine ecosystems in the sea.