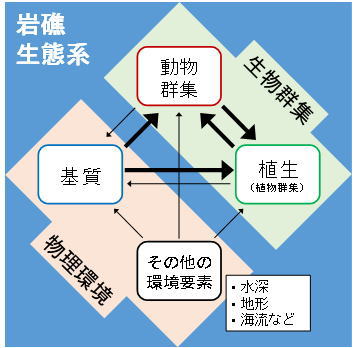
このように複雑な生物群集を有する岩礁生態系ですが、特に「植生」と「基質」という環境の要素によってその状態は大きく左右されます。これら2つの要素が生物群集にどのような影響を与えるのか、それぞれ考えてみることにしましょう。
植生とは、その場所にどんな植物(岩礁域の場合は海藻類)がどんな量・割合で生えているかを表す言葉で、生物群集中で植物の部分だけを見たものともみなせます。岩礁域における植生の典型例は、アラメ(Eisenia bicyclis)やカジメ(Ecklonia cava)等の多年生の大型海藻が繁茂する、いわゆる海中林と呼ばれるものです。海中林は衰退と回復を一年の間に繰り返すとされますが、様々な原因から衰退状態が長期間続いてしまうことがあります1。海中林に住む底生動物群集は、主要な一次生産者である大型海藻を起点とする食物連鎖から成り立っているため(図1)、海中林が消失すると大きな構造変化を生じます。とりわけ、餌として大型海藻に強く依存するアワビ類等の植食性動物は急激な減少を示します(図2)。
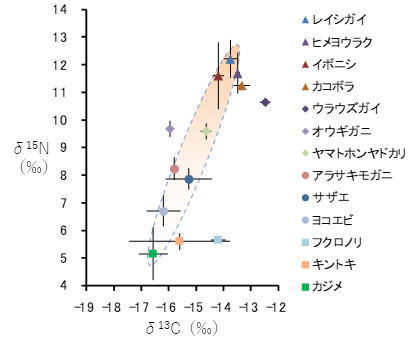
右上ほど栄養段階が高いことを表す。同一の食物連鎖を構成する種は右上がりの直線状に並ぶ。
岩礁の生態系には、岩盤や礫・転石等、海底面を構成する基質の特性も大きく影響します4。アワビ等の増殖を目的として作られた岩礁(人工礁)の多くは、沈められた礫同士ががっちりと組み合わさり、波などで簡単には動かない基質から成っています(図3)。

人為的に沈められた礫が組み合ってほとんど動かない状態になっている。
ここまでをまとめると、岩礁生態系の概念図は図5のようになります。こうした要素それぞれの働きを理解することは、岩礁生態系の管理を考える上でも大切です。
底生動物群集を定量的に調べる代表的な方法に、枠取り調査と呼ばれるものがあります。これは、大きさの決まった正方形の枠を、対象とする岩礁域内にランダムに複数設置し、枠の中の底生動物を全て採集するという調査です。潮下帯の枠取り調査では空気ボンベを背負って潜水する必要がありますが、潮間帯でも同様の方法で調査を行うことができます(図6)。

こうして採集された底生動物のサンプルから、種数・種組成やバイオマス等の群集に関する重要なデータを得ることができます。なお、このような生物相調査は、一回きりではなく定期的に行うことで大きな効果を発揮します。過去のデータの季節変化や経年変化を把握することは、現在の岩礁生態系がどのような経過をたどって今の状態となり、また今後どのような状態に向かっていくのか推測するための材料になるからです。
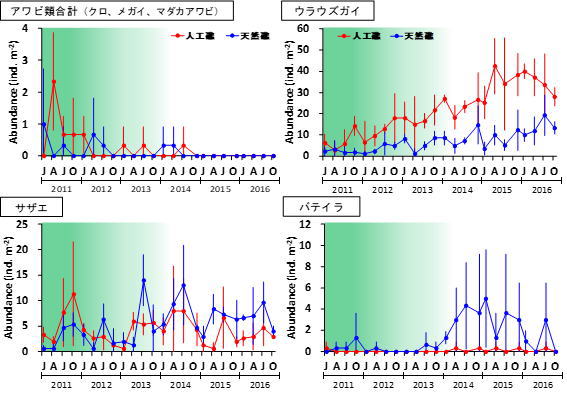
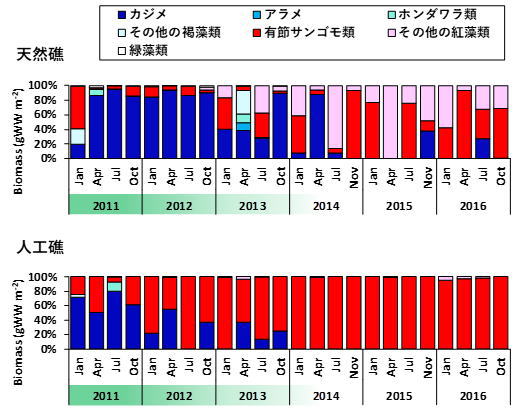
例として、中央水産研究所横須賀庁舎が長期にわたり実施してきた生物相調査5の結果を一部ご紹介します。神奈川県三浦半島西部に位置する長井地先の水深約12 mの海底には、2008年にアワビ等の磯根資源の増殖・保護のための人工礁が設置されました。この調査は、設置後の人工礁と周囲の天然礁における生態系の状態の変化を調べるために行われてきたものです。人工礁と天然礁では、2013年ごろまではカジメを主体とする海中林が形成されていましたが、2014年以降海中林が衰退し、現在は完全に消失した状態にあります(図4)。
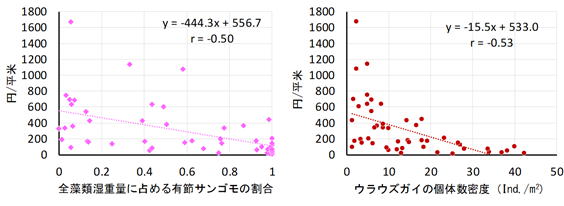
底生動物の種数を見ると、海中林が消失する前後で大きな変化はありませんでした。しかし、群集中の種構成は2014年を境に変化していることが分かりました。例えば、アワビ類などの水産上重要な種は、海中林が消失した後に個体数密度が低下していました。しかし、同じく海藻を餌とするサザエやバテイラ(Omphalius pfeifferi pfeifferi)については、天然礁でのみ増加傾向が見られました(図2)。これらの種はアワビ類ほど大型藻類への依存性が高くないため、他の小型海藻を餌として利用することができたものと考えられます。一方で、基質の安定性が高い人工礁では有節サンゴモの圧倒的な優占状態となったため、サザエやバテイラの餌となるような小型海藻も少なく、これらの種の増加が抑制された可能性があります。その代わり、人工礁ではウラウズガイ(Astralium haematragum)という小型の巻貝が増加し、最優占種となっていました(図2)。この現象は、海中林消失後の人工礁が、ウラウズガイにとって生き残りに適した環境であったために起きたと考えられます(ウラウズガイ増加の謎を生態から考える)。多様度に全体の個体数をかけ合わせた「群集繁栄度」という概念で比較すると、優占種の影響を大きく見た場合ほど、海中林消失後の礁間の差が大きくなっていました(図7)。つまり、海中林が消失したことで人工礁に生息する種の中で個体数の偏りが大きくなったことが、礁間の群集構造の違いをもたらしたと考えられます。多様度や群集繁栄度の概念についての解説や、群集解析の具体的な方法については、こちらのリンクをご覧ください(森下の群集繁栄度)。このように、海中林が消失した際に底生動物群集がどのように応答するかは、人工礁と天然礁では異なることが分かりました。
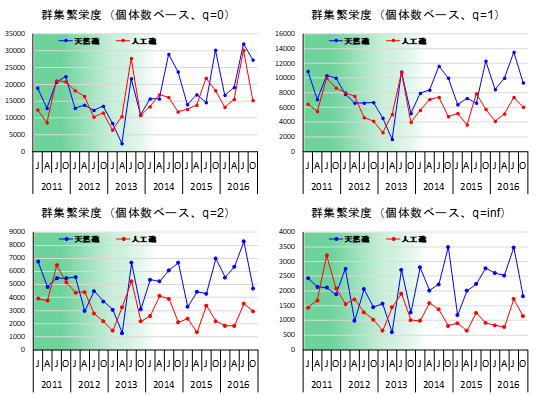
感度パラメータqの値が大きいほどその群集が含む優占種の与える負の影響が大きくなる。
緑色は海中林が形成されていた時期。
岩礁域の生態系が我われ人間にとって望ましくない状態に向かっているとしたら、そのことをいち早く把握することが資源の安定的・持続的な利用の上で重要です。海中林の衰退した状態が長期間続くと、岩礁域の水産上の価値は低下し、経済的にも大きな悪影響が出てしまいます(図8)。これは、経済価値の特に高いアワビ類が大きく減少すること、ウニ類の身入りが悪くなり経済価値が無くなること等が原因です。底生動物への影響以外にも、稚魚の生育場やイカ類の産卵場が減少する等の間接的な影響も生じるため、資源の安定利用の上で大きな問題となります。一般的に、海中林が衰退すると生物多様性が下がると言われますが、種数や多様度の値に大きな影響が現れるとは限りません。海中林の衰退に伴って生態系がどのように変化しているか把握するには、優占種の与える影響の度合いを変えて群集繁栄度を比較するのが効果的です(図7)。
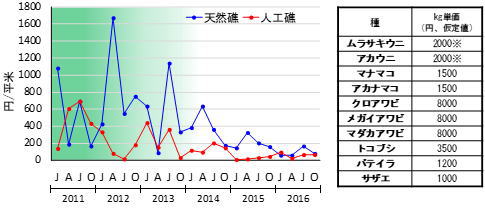
のkg単価仮定値(円、右表参照)と枠取り調査で得られた1 m2当たりの湿重量(kg)をかけ合わせ
て計算。ムラサキウニとアカウニは海中林が衰退すると身入りの悪化により無価値化するため、
2011~13年は2000円/kg、2014~16年は0円/kgとして計算。
また、近年では底生動物に加えてアイゴ等の植食性魚類が与える影響も大きいと考えられるため(図10)、刺し網等を利用した大型海藻の防護も対策となります。
水中カメラによる魚類の行動の定点観察の手法については、こちらのリンクをご覧ください(カメラで調べる魚類の多様性)。

上に述べたように、岩礁域の資源の安定的かつ持続的な利用のためには、生態系管理を適切に行う必要があります。このように人の手を加えることで生産性が維持された沿岸海域は「里海」とも呼ばれ6、その意義がだんだんと認知されてきています。なお、今後は温暖化など地球規模の環境変化が進行することによる沿岸域への影響も危惧されています。海水温の上昇は、海中林の衰退のほか、新たな生物の移入や既存の種の絶滅などを引き起こし、結果として沿岸岩礁域の生産性を低下させる恐れがあります。こうした大きな変化に対して順応的に管理方策を考えるためにも、継続的な生物相調査を行うことで生態系の変化の過程をモニタリングしていくことの重要性は高まっていると言えます。
2. Hayakawa, J., Kawamura, T., Ohashi, S., Horii, T., & Watanabe, Y. (2008) Habitat selection of Japanese top shell (Turbo cornutus) on articulated coralline algae; combination of preferences in settlement and post-settlement stage. Journal of Experimental Marine Biology and Ecology, 363(1), 118-123.
3. Hayakawa, J., Kawamura, T., Kurogi, H., & Watanabe, Y. (2013) Shelter effects of coralline algal turfs: protection for Turbo cornutus juveniles from predation by a predatory gastropod and wrasse. Fisheries science, 79(1), 15-20.
4. 川俣茂(1994)磯根漁場造成における物理的攪乱の重要性.水産工学, 31(2), 103-110.
5. 丹羽健太郎・寺本航・黒木洋明(2016)浅海域の造成漁場と天然礁における底生生物相
(特集 沿岸漁場における生物多様性). 海洋と生物, 38(6), 667-674.
6.柳哲雄(2006)「里海論」恒星社厚生閣,東京.